
微量轉錄組測序揭示TET酶在動物卵母細胞成熟和孤雌胚胎發育中的作用
易小結
單細胞組學技術為探明哺乳動物卵母細胞早期生長分子機制提供了新機遇。此前,單細胞DNA甲基化(scBS-seq)與轉錄組測序(Smart-seq2)已揭示豬生發泡卵母細胞成熟的關鍵調控機制,闡明了減數分裂停止到恢復的分子過程(詳見:項目文章 | 單細胞DNA甲基化與轉錄組分析揭示豬生發泡卵母細胞成熟的關鍵調控機制)。TET家族酶在基因組去甲基化中扮演重要角色,對胚胎發育和干細胞多能性至關重要,但在豬卵母細胞中的具體功能仍需深入探究。
本研究發現TET家族酶(TET1、TET2、TET3)在豬卵母細胞成熟和孤雌胚胎發育中發揮關鍵作用。Bobcat339處理通過改變5mC和5hmC水平,顯著影響卵母細胞成熟、紡錘體結構、染色體排列及胚胎發育進程。此外,TET酶還通過調控合子基因組激活(ZGA)及多能性相關基因的表達,進一步影響胚胎發育。這些發現為理解TET家族在哺乳動物胚胎發育中的功能提供了新視角。
Smart-seq2技術是本研究的核心支持,其高通量轉錄組分析揭示了Bobcat339處理后胚胎的差異表達基因及發育停滯的分子機制。該技術為深入理解胚胎發育分子過程提供了重要線索和數據支持,為后續基因功能驗證及豬生殖生物學和胚胎工程研究奠定了堅實的理論基礎。
研究摘要
近日,湖北省農業科學院畜牧獸醫研究所陳礬博士(助理研究員)為第一作者,黃濤副研究員和畢延震研究員為共同通訊,在《International Journal of Molecular Sciences》(Int. J. Mol. Sci. )期刊發表題為《TET Family Members Are Integral to Porcine Oocyte Maturation and Parthenogenetic Pre-Implantation Embryogenesis》的科研成果。研究通過使用Bobcat339(一種特異性的小分子TET家族抑制劑)處理以抑制TET酶活性,進而觀察其對豬卵母細胞成熟和早期胚胎發育的影響。研究結果揭示了TET家族酶在調控豬卵母細胞成熟、紡錘體結構、染色體排列以及胚胎發育過程中5mC和5hmC水平的關鍵作用。研究闡明了TET酶在豬卵母細胞成熟和孤雌胚胎發育中的具體作用機制,為理解哺乳動物胚胎發育的表觀遺傳調控提供新的視角。此外研究結果也為豬的生殖生物學和胚胎工程提供重要理論基礎,有助于優化豬的繁殖技術和胚胎發育調控策略。

標題:TET Family Members Are Integral to Porcine Oocyte Maturation and Parthenogenetic Pre-Implantation Embryogenesis(TET家族成員對豬卵母細胞成熟和孤雌胚胎發育至關重要)
發表時間:2023-08-05
發表期刊:Int. J. Mol. Sci.
影響因子:IF 5.6/Q2
技術平臺:Smart-seq2等
研究方法
樣本收集:從雌豬卵巢中分離3-6 mm的卵泡,收集帶有均勻細胞質和多層卵丘細胞的卵丘卵母細胞復合體(cumulus oocyte complexes, COCs)。
體外成熟(IVM):將COCs在培養基中培養,添加促性腺激素(eCG和hCG)以促進卵母細胞成熟。
孤雌激活:去除卵丘細胞后,激活卵母細胞,激活后的卵母細胞在培養基中培養。
Bobcat339處理:將Bobcat339溶解于DMSO中,稀釋至不同濃度(0μM、50μM、100μM、200μM、400μM),添加到成熟培養基中以抑制TET酶活性。
檢測方法:
免疫熒光染色:檢測5mC和5hmC水平。
qRT-PCR:用于檢測基因表達水平。
微量RNA測序(Smart-seq2):分析胚胎轉錄組的變化。
結果圖形
(1)Bobcat339處理阻斷豬卵母細胞第一極體(First Polar Body)排出
實驗中,不同濃度的Bobcat339處理顯著降低了豬卵母細胞第一極體排出率(PBE),表現為濃度依賴性。在200μM和400μM濃度下,PBE率顯著下降,表明Bobcat339抑制TET酶活性后,卵母細胞成熟過程受到阻礙。
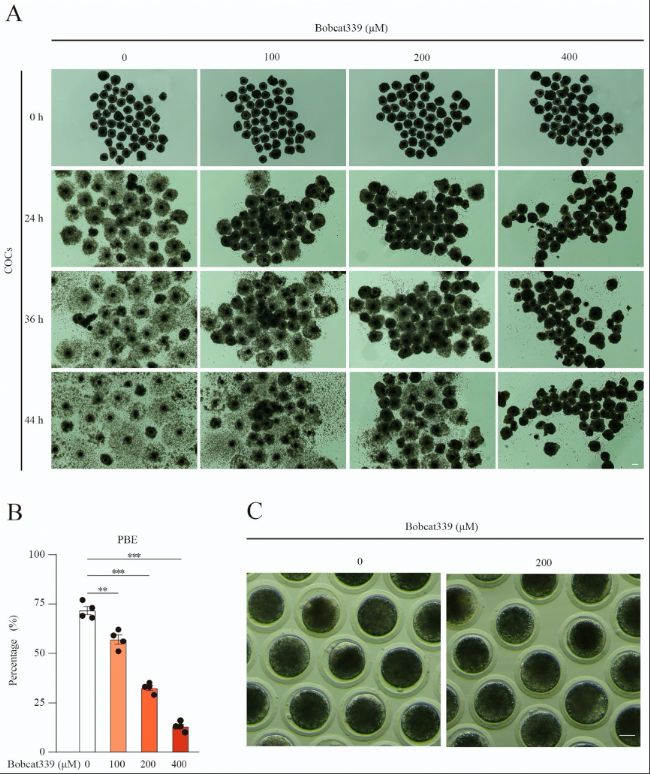
(B)經100μM、200μM和400μM Bobcat339處理后,第一極體排出(PBE)率顯著降低。
(C)對照組和經200μM Bobcat339處理的卵母細胞在體外培養44小時后的第一極體排出的代表性圖像。
(2)經Bobcat339處理引發豬卵母細胞凋亡
通過Annexin-V-FITC染色檢測早期凋亡水平,發現Bobcat339處理后的卵母細胞早期凋亡率顯著增加。此外,BAX和BCL-2的mRNA水平顯著升高,BAX/BCL-2比值顯著上升,表明TET酶抑制導致卵母細胞早期凋亡。
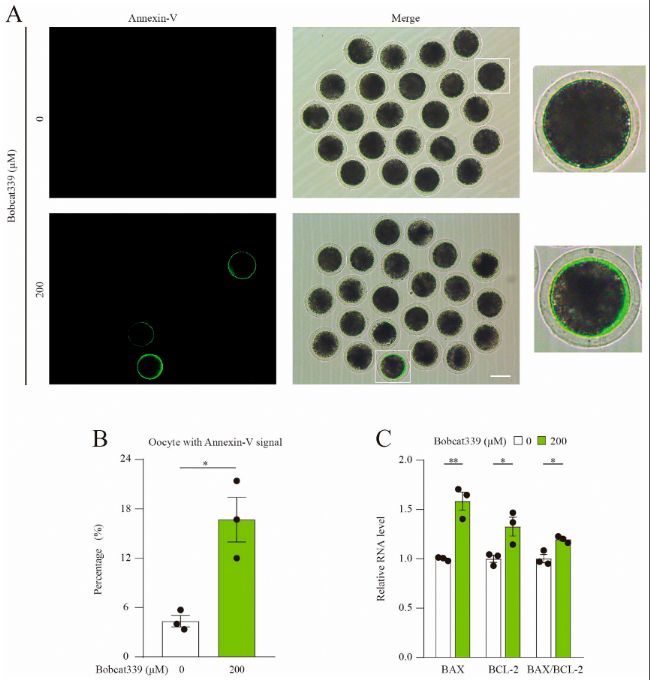
(B)對照組和經Bobcat339處理的卵母細胞中帶有Annexin-V信號的卵母細胞比例。
(C)qRT-PCR檢測對照組和經Bobcat339處理的卵母細胞BAX、BCL-2、BAX/BCL-2的mRNA水平。
(3)Bobcat339對卵母細胞紡錘體結構和染色體排列的作用
使用α-tubulin染色觀察紡錘體形態,結果顯示Bobcat339處理的卵母細胞紡中錘體結構異常,染色體排列分散。與對照組相比,異常紡錘體和染色體排列比例顯著增加,表明TET酶對卵母細胞的紡錘體結構和染色體排列具有重要作用。
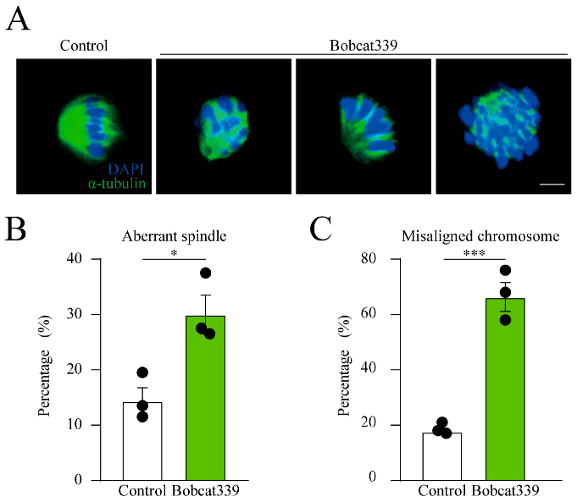
(B)經Bobcat339處理的卵母細胞中異常紡錘體比例顯著增加。
(C)經Bobcat339處理的卵母細胞中染色體錯位比例顯著升高。
(4)Bobcat339處理導致豬卵母細胞中的5mC/5hmC水平發生變化
免疫熒光染色結果顯示,Bobcat339處理的卵母細胞中5hmC信號顯著降低,而5mC信號顯著增加。定量分析進一步驗證這一結果,表明TET酶通過調節5mC和5hmC水平影響卵母細胞成熟。
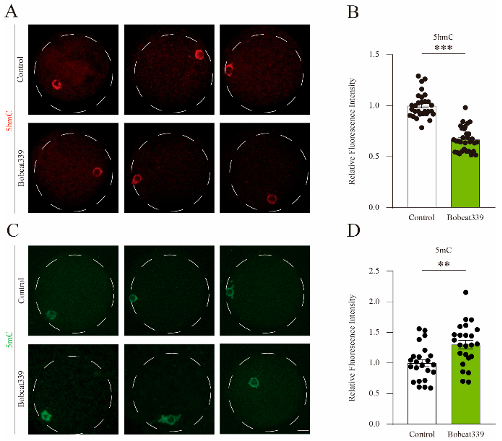
(B)對照組和經Bobcat339處理的卵母細胞中5hmC的熒光強度。
(C)對照組和經Bobcat339處理的卵母細胞中5mC的免疫熒光染色。
(D)對照組和經Bobcat339處理的卵母細胞中5mC的平均熒光強度。
(5)Bobcat339對胚胎發育的影響
在孤雌胚胎發育實驗中,不同濃度的Bobcat339處理顯著降低了胚胎發育到囊胚階段的比例。胚胎在4細胞階段停滯,表明Bobcat339抑制TET酶活性后,胚胎發育受到嚴重影響。
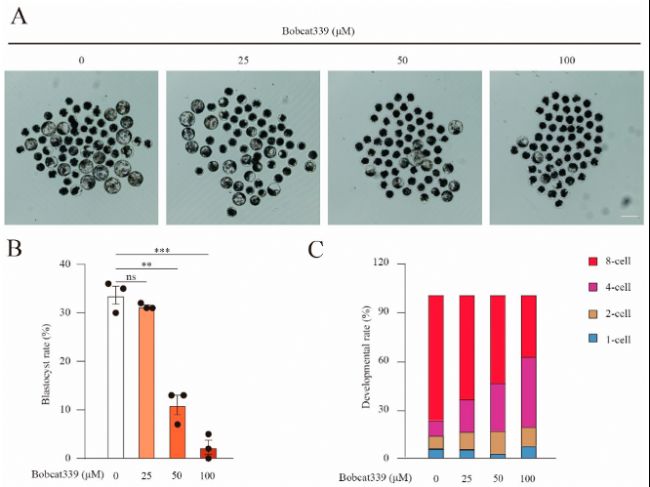
圖5:Bobcat339處理抑制了豬的早期胚胎發育。
(A)對照組和經Bobcat339處理(25、50和100 μM)的孤雌激活豬胚胎在第6天囊胚形成的圖像。
(B)對照組和經Bobcat339處理組中發育到囊胚階段的激活卵母細胞比例。
(C)對照組和經Bobcat339處理(25、50和100 μM Bobcat339)組中第3天不同階段的胚胎比例。
(6)Bobcat339處理降低了ZGA和多能性相關基因的表達
qRT-PCR檢測結果顯示,Bobcat339處理后,合子基因組激活(zygotic gene activation,ZGA)標記基因(如EIF1A和DPPA2)以及多能性相關基因(如OCT4和NANOG)的mRNA水平顯著下降,表明Bobcat339處理破壞了ZGA,影響胚胎發育。
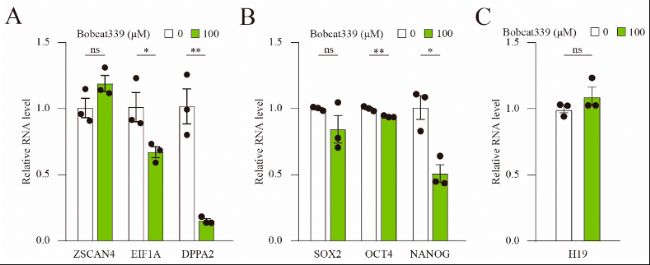
圖6:Bobcat339處理阻斷了ZGA和多能性基因的表達。
(A)經Bobcat339處理的胚胎中,EIF1A和DPPA2的mRNA水平顯著降低。
(B)經Bobcat339處理的胚胎中,OCT4和NANOG的mRNA水平降低;SOX2的mRNA水平則沒有變化。
(C)Bobcat339處理后,H19的mRNA水平未受影響。
(7)Bobcat339處理影響豬胚胎中的5mC/5hmC水平
免疫熒光染色和定量分析顯示,Bobcat339處理的胚胎中5hmC水平顯著降低,而5mC水平顯著增加,表明TET家族酶通過調節5mC和5hmC水平影響胚胎發育。
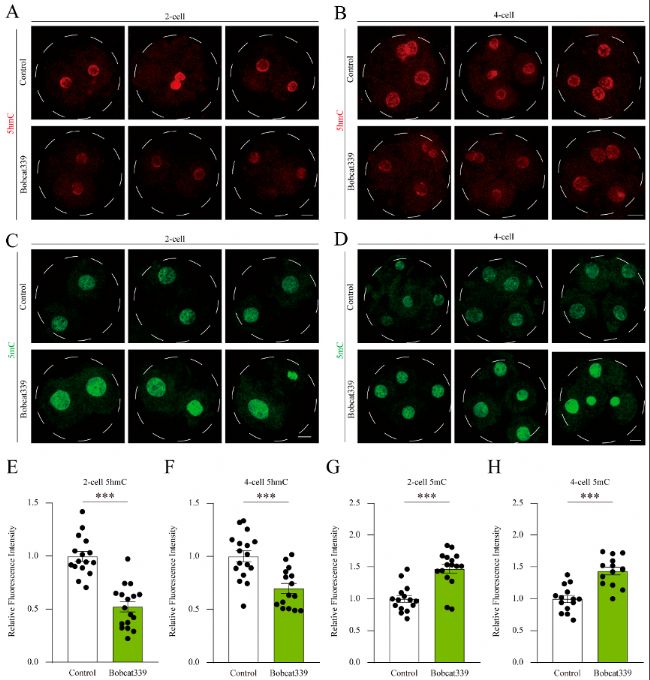
圖7:Bobcat339處理影響豬胚胎中的5mC和5hmC水平。
(A-B) 對照組和經Bobcat339處理的胚胎在2細胞和4細胞階段的5hmC免疫熒光染色。
(C-D) 對照組和經Bobcat339處理的胚胎在2細胞和4細胞階段的5mC免疫熒光染色。
(E-F) 在對照組和經Bobcat339處理的胚胎中,2細胞和4細胞階段的5hmC熒光強度。
(G-H) 在對照組和經Bobcat339處理的胚胎中,2細胞和4細胞階段的5mC平均熒光強度。
(8)微量轉錄組數據的比較分析
通過Smart-seq2技術對4細胞階段的胚胎進行RNA測序分析,分析結果鑒定出Bobcat339處理后胚胎中的203個差異表達基因(DEGs),其中75個上調基因,128個下調基因。基因本體(GO)分析顯示,這些差異表達基因主要富集在細胞增殖、細胞組分與線粒體相關以及細胞黏附分子結合等生物學過程中。
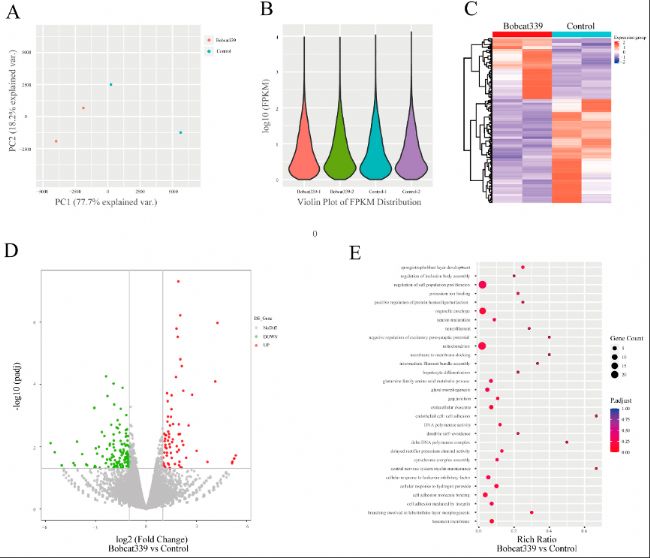
圖8:對照組和經Bobcat339處理的胚胎之間的差異基因表達模式。
(A)對照組和經Bobcat339處理的胚胎在4細胞階段的mRNA水平的主成分分析(PCA)。
(B)對照組和經Bobcat339處理組之間的基因表達和分布的小提琴圖。
(C)對照組和經Bobcat339處理的胚胎在4細胞階段的差異表達基因(DEGs)的分層聚類。
(D)對照組和經Bobcat339處理的胚胎在4細胞階段DEGs火山圖。紅色(上調)/綠色(下調)。
(E)基因本體(GO)分析顯示DEGs在某些生物學過程、分子功能和細胞組分中富集。
參考文獻:
Chen, F.; Li, M.-G.; Hua, Z.-D.; Ren, H.-Y.; Gu, H.; Luo, A.-F.; Zhou, C.-F.; Zhu, Z.; Huang, T.; Bi, Y.-Z. TET Family Members Are Integral to Porcine Oocyte Maturation and Parthenogenetic Pre-Implantation Embryogenesis. Int. J. Mol. Sci. 2023, 24, 12455. https://doi.org/10.3390/ijms241512455.
單細胞組學技術為探明哺乳動物卵母細胞早期生長分子機制提供了新機遇。此前,單細胞DNA甲基化(scBS-seq)與轉錄組測序(Smart-seq2)已揭示豬生發泡卵母細胞成熟的關鍵調控機制,闡明了減數分裂停止到恢復的分子過程(詳見:項目文章 | 單細胞DNA甲基化與轉錄組分析揭示豬生發泡卵母細胞成熟的關鍵調控機制)。TET家族酶在基因組去甲基化中扮演重要角色,對胚胎發育和干細胞多能性至關重要,但在豬卵母細胞中的具體功能仍需深入探究。
本研究發現TET家族酶(TET1、TET2、TET3)在豬卵母細胞成熟和孤雌胚胎發育中發揮關鍵作用。Bobcat339處理通過改變5mC和5hmC水平,顯著影響卵母細胞成熟、紡錘體結構、染色體排列及胚胎發育進程。此外,TET酶還通過調控合子基因組激活(ZGA)及多能性相關基因的表達,進一步影響胚胎發育。這些發現為理解TET家族在哺乳動物胚胎發育中的功能提供了新視角。
Smart-seq2技術是本研究的核心支持,其高通量轉錄組分析揭示了Bobcat339處理后胚胎的差異表達基因及發育停滯的分子機制。該技術為深入理解胚胎發育分子過程提供了重要線索和數據支持,為后續基因功能驗證及豬生殖生物學和胚胎工程研究奠定了堅實的理論基礎。
研究摘要
近日,湖北省農業科學院畜牧獸醫研究所陳礬博士(助理研究員)為第一作者,黃濤副研究員和畢延震研究員為共同通訊,在《International Journal of Molecular Sciences》(Int. J. Mol. Sci. )期刊發表題為《TET Family Members Are Integral to Porcine Oocyte Maturation and Parthenogenetic Pre-Implantation Embryogenesis》的科研成果。研究通過使用Bobcat339(一種特異性的小分子TET家族抑制劑)處理以抑制TET酶活性,進而觀察其對豬卵母細胞成熟和早期胚胎發育的影響。研究結果揭示了TET家族酶在調控豬卵母細胞成熟、紡錘體結構、染色體排列以及胚胎發育過程中5mC和5hmC水平的關鍵作用。研究闡明了TET酶在豬卵母細胞成熟和孤雌胚胎發育中的具體作用機制,為理解哺乳動物胚胎發育的表觀遺傳調控提供新的視角。此外研究結果也為豬的生殖生物學和胚胎工程提供重要理論基礎,有助于優化豬的繁殖技術和胚胎發育調控策略。
標題:TET Family Members Are Integral to Porcine Oocyte Maturation and Parthenogenetic Pre-Implantation Embryogenesis(TET家族成員對豬卵母細胞成熟和孤雌胚胎發育至關重要)
發表時間:2023-08-05
發表期刊:Int. J. Mol. Sci.
影響因子:IF 5.6/Q2
技術平臺:Smart-seq2等
研究方法
樣本收集:從雌豬卵巢中分離3-6 mm的卵泡,收集帶有均勻細胞質和多層卵丘細胞的卵丘卵母細胞復合體(cumulus oocyte complexes, COCs)。
體外成熟(IVM):將COCs在培養基中培養,添加促性腺激素(eCG和hCG)以促進卵母細胞成熟。
孤雌激活:去除卵丘細胞后,激活卵母細胞,激活后的卵母細胞在培養基中培養。
Bobcat339處理:將Bobcat339溶解于DMSO中,稀釋至不同濃度(0μM、50μM、100μM、200μM、400μM),添加到成熟培養基中以抑制TET酶活性。
檢測方法:
免疫熒光染色:檢測5mC和5hmC水平。
qRT-PCR:用于檢測基因表達水平。
微量RNA測序(Smart-seq2):分析胚胎轉錄組的變化。
結果圖形
(1)Bobcat339處理阻斷豬卵母細胞第一極體(First Polar Body)排出
實驗中,不同濃度的Bobcat339處理顯著降低了豬卵母細胞第一極體排出率(PBE),表現為濃度依賴性。在200μM和400μM濃度下,PBE率顯著下降,表明Bobcat339抑制TET酶活性后,卵母細胞成熟過程受到阻礙。
圖1:Bobcat339處理抑制了豬卵母細胞成熟。
(A)對照組和經Bobcat339處理(0、100、200和400 μM)的卵丘卵母細胞復合體(COCs)在體外培養24、36和44小時后的代表性圖像。對照組的卵丘細胞擴張良好,而Bobcat339處理組的卵丘細胞擴張較少且更加黏附。(B)經100μM、200μM和400μM Bobcat339處理后,第一極體排出(PBE)率顯著降低。
(C)對照組和經200μM Bobcat339處理的卵母細胞在體外培養44小時后的第一極體排出的代表性圖像。
(2)經Bobcat339處理引發豬卵母細胞凋亡
通過Annexin-V-FITC染色檢測早期凋亡水平,發現Bobcat339處理后的卵母細胞早期凋亡率顯著增加。此外,BAX和BCL-2的mRNA水平顯著升高,BAX/BCL-2比值顯著上升,表明TET酶抑制導致卵母細胞早期凋亡。
圖2:Bobcat339處理誘導豬卵母細胞早期凋亡。
(A)對照組和經Bobcat339處理的卵母細胞的早期凋亡熒光信號。(B)對照組和經Bobcat339處理的卵母細胞中帶有Annexin-V信號的卵母細胞比例。
(C)qRT-PCR檢測對照組和經Bobcat339處理的卵母細胞BAX、BCL-2、BAX/BCL-2的mRNA水平。
(3)Bobcat339對卵母細胞紡錘體結構和染色體排列的作用
使用α-tubulin染色觀察紡錘體形態,結果顯示Bobcat339處理的卵母細胞紡中錘體結構異常,染色體排列分散。與對照組相比,異常紡錘體和染色體排列比例顯著增加,表明TET酶對卵母細胞的紡錘體結構和染色體排列具有重要作用。
圖3:Bobcat339對紡錘體形態和染色體排列的影響。
(A)對照組和經Bobcat339處理的卵母細胞(200μM Bobcat339)的紡錘體形態和染色體排列圖像。(B)經Bobcat339處理的卵母細胞中異常紡錘體比例顯著增加。
(C)經Bobcat339處理的卵母細胞中染色體錯位比例顯著升高。
(4)Bobcat339處理導致豬卵母細胞中的5mC/5hmC水平發生變化
免疫熒光染色結果顯示,Bobcat339處理的卵母細胞中5hmC信號顯著降低,而5mC信號顯著增加。定量分析進一步驗證這一結果,表明TET酶通過調節5mC和5hmC水平影響卵母細胞成熟。
圖4:Bobcat339處理改變了豬卵母細胞中的5mC和5hmC水平。
(A)豬卵母細胞中5hmC的免疫熒光染色。(B)對照組和經Bobcat339處理的卵母細胞中5hmC的熒光強度。
(C)對照組和經Bobcat339處理的卵母細胞中5mC的免疫熒光染色。
(D)對照組和經Bobcat339處理的卵母細胞中5mC的平均熒光強度。
(5)Bobcat339對胚胎發育的影響
在孤雌胚胎發育實驗中,不同濃度的Bobcat339處理顯著降低了胚胎發育到囊胚階段的比例。胚胎在4細胞階段停滯,表明Bobcat339抑制TET酶活性后,胚胎發育受到嚴重影響。
圖5:Bobcat339處理抑制了豬的早期胚胎發育。
(B)對照組和經Bobcat339處理組中發育到囊胚階段的激活卵母細胞比例。
(C)對照組和經Bobcat339處理(25、50和100 μM Bobcat339)組中第3天不同階段的胚胎比例。
(6)Bobcat339處理降低了ZGA和多能性相關基因的表達
qRT-PCR檢測結果顯示,Bobcat339處理后,合子基因組激活(zygotic gene activation,ZGA)標記基因(如EIF1A和DPPA2)以及多能性相關基因(如OCT4和NANOG)的mRNA水平顯著下降,表明Bobcat339處理破壞了ZGA,影響胚胎發育。
圖6:Bobcat339處理阻斷了ZGA和多能性基因的表達。
(B)經Bobcat339處理的胚胎中,OCT4和NANOG的mRNA水平降低;SOX2的mRNA水平則沒有變化。
(C)Bobcat339處理后,H19的mRNA水平未受影響。
(7)Bobcat339處理影響豬胚胎中的5mC/5hmC水平
免疫熒光染色和定量分析顯示,Bobcat339處理的胚胎中5hmC水平顯著降低,而5mC水平顯著增加,表明TET家族酶通過調節5mC和5hmC水平影響胚胎發育。
圖7:Bobcat339處理影響豬胚胎中的5mC和5hmC水平。
(C-D) 對照組和經Bobcat339處理的胚胎在2細胞和4細胞階段的5mC免疫熒光染色。
(E-F) 在對照組和經Bobcat339處理的胚胎中,2細胞和4細胞階段的5hmC熒光強度。
(G-H) 在對照組和經Bobcat339處理的胚胎中,2細胞和4細胞階段的5mC平均熒光強度。
(8)微量轉錄組數據的比較分析
通過Smart-seq2技術對4細胞階段的胚胎進行RNA測序分析,分析結果鑒定出Bobcat339處理后胚胎中的203個差異表達基因(DEGs),其中75個上調基因,128個下調基因。基因本體(GO)分析顯示,這些差異表達基因主要富集在細胞增殖、細胞組分與線粒體相關以及細胞黏附分子結合等生物學過程中。
圖8:對照組和經Bobcat339處理的胚胎之間的差異基因表達模式。
(B)對照組和經Bobcat339處理組之間的基因表達和分布的小提琴圖。
(C)對照組和經Bobcat339處理的胚胎在4細胞階段的差異表達基因(DEGs)的分層聚類。
(D)對照組和經Bobcat339處理的胚胎在4細胞階段DEGs火山圖。紅色(上調)/綠色(下調)。
(E)基因本體(GO)分析顯示DEGs在某些生物學過程、分子功能和細胞組分中富集。
參考文獻:
Chen, F.; Li, M.-G.; Hua, Z.-D.; Ren, H.-Y.; Gu, H.; Luo, A.-F.; Zhou, C.-F.; Zhu, Z.; Huang, T.; Bi, Y.-Z. TET Family Members Are Integral to Porcine Oocyte Maturation and Parthenogenetic Pre-Implantation Embryogenesis. Int. J. Mol. Sci. 2023, 24, 12455. https://doi.org/10.3390/ijms241512455.
標簽:
DNA甲基化



Copyright(C) 1998-2025 生物器材網 電話:021-64166852;13621656896 E-mail:info@bio-equip.com